
�֤ΰ�����ͭ���֤ΰ���
��C.dowiana var.
aurea��Stella�ǤΥ���ո���βֿ���

�Ϥ����
ɮ�Ԥ�����Ϲ���Ū�ˤϡز��á٤Ǥ��뤬������Ū�ˤϡإ��ʿ�ʪ�٤Ǥ��롣����ˤ�ä�����Ūʬ��ˤĤ��ƿҤͤ���ȡ��ؿ�ʪ�Х����ƥ��Υ������٤����翦�������Ƥ��ޤ����ܿ��ϥХ��ƥ����õ�ῴ�����溲���Ż����ȻפäƤ���Τ����������ȤƲԤ��ʤ���Фʤ�ʤ��Τǡ���������������ܲ�Ҥ�Ĥ�Ǥ��롣�����餤�Ĥ�����ع�������ζ��ܤФ����ȤäƤ���櫓�Ǥ�ʤ����ޤ�˿��������Ȥ˽б�Ф��ꤷ�Ƥ���櫓�Ǥ�ʤ������Ÿ�ǥ������䤷�Ƥ���Ȥ��⤢��С��������Ϥ������͢�������ꡢ���������ȥե饹���Ĥ��������Ƥ���Ȥ��⤢�롣
���ޤ��飴ǯ���ˤ���ե饹���Ĥ������������䤷�����Ȥ����ä�������Ϥ��Υե饹���Ĥ����֤ˤ����ꡢ���βֿ����Ѳ��ˤĤ��Ƥ������������������Ƥ����Τǡ��ֿ��ΰ����˴ؤ����ä���Ϫ���Ƥߤ������������˴�ñ�ʰ����˴ؤ����ä�����⤷�Ƥ����ʤ���Фʤ�ʤ�������ˡּ�ȤϤʤˤ����פȤ�������ʥơ��ޤˤĤ��ơ�����ϸ�äƤ����ʤ���Фʤ�ʤ��褦����
���ְ����ؤδ���
���زֿ��ΰ����˴ؤ����á٤ο�������������äƤ��ʤ��������ǣ�ǯ�����ܻ�ˡֹ���βֿ��˴ؤ�������ءפ����⤵��Ƥ����Τ����ä��Τǡ��������Ѥ��Ʋֿ��ΰ����ˤĤ��Ʋ�����ߤ���������������ᤫ�龵���ϼ����Ƥ���Τǡ��ɤ��ޤǴ�ñ�˲���Ǥ��뤫�狼��ʤ������Ȥˤ����ä�ʤ�Ƥ�������
ȴ������
�����٤Ƥ�1909ǯ�ѹ�����ؼԤǤ���Hurst��Τθ��椫��Ϥޤä�������ϸ��۰��η�̤�����ʿ�ʪ����֤ΰ����ͼ�����Ϥ�����������ΤǤ��ä���
��������������С��إ��ʿ�ʪ�����Ʊ�Τθ��ۤˤ����Ƥ⡢2����ΰ����Ҥ�ξ�������ä��Ȥ��ˡ��ȿ��βֿ���������٤Ȥ������ȤǤ��롣2����ΰ����ҤȤϹȿ����Ǥ�����ʪ����¸�ݤ�ɽ�������ҡȣá���ɡ������̵��������ʪ�����ȿ����Ǥ��������뵡���˴�Ϳ��������ҡȣҡ���ɤǤ������ȿ���������
�����ߤޤǤˡ��͡��ʺ�ʪ�Ǹ¸��������֤�����ʰʳ��Ǥ�Ʊ�ͤʸ��ݤ���ǧ����Ƥ��롣�Ĥޤ����֤��Ф��褦�Ȥ��ơ����Ʊ�Τ���碌����Ȳ֤ˤʤäƤ��ޤä��٤Ȥ������ݤǤ��롣�������ȥԡ������Ʊ�Τ����ȡ����줫��Ҳ𤹤������ɤ��ꡢ�ƣ��ʻ����裱��ˤˤϤ��٤ƻ�β֤�餫���뤳�ȤȤʤ롣�����Ƥ��λ��Ʊ�ΤΤ������碌�λҶ��ʣƣ��������裲��ˤϻ硧���9:7����Ψ�Ȥʤ�Τ�����
���ʾ�����Ƥ�����Ȳֿ��ΰ����ϿƤ��Ѥ�������ҷ��ˤ�äơ���������פ�ֹȡ߹ȡ���פˤʤäƤ��ޤ��餷���������礭�ʸ����Ȥ��ơ��Ȳ֤βֿ���ȯ�����뤿��ˤϡزֿ��κ����٤ȡ�ȯ�������륷���ƥ�٤�·��ʤ��ƤϤʤ�ʤ��餷�����ɤ��餫�����Ǥ�礱�Ƥ��ޤ���ȯ���Ǥ�������֤ȤʤäƤ��ޤ��Τ�����ɮ�Ԥ���غ߳���ˡ������ؤιֵ�������Ƥ����������ϵ���û���ä�����������������Ω�Ƥʤ���Τ��ȴ����Ƥ������Ȥ��ݤ�ʤ������������ºݤ˸��ۤˤ�äƿ��ʼ���Ф����Ǥϡ��ΤäƤ������ۤ����褤�μ���¿���Τǡ����ε���˰����ؤδ�ñ�ʴ��ä����Ǥ�ؤ�Ǥߤ�Τ�褤�Ǥ�������
�����줫����⤷�Ƥ����˻Ϥޤꡢ��ǧ���Ƥ�����ĺ���������Ȥ����롣����ϻ����δ��ʸ���Ф�����ɾ���Ƥ���Ω��塢�֡������餷���פ�ɽ������٤��ʤΤ��������Ρ֡������餷���פȤ���ɽ��ˡ�Ϥ鷺��路���Τǰʸ��ά���뤳�Ȥˤ�������λ��ĺ������������˻����ˤϿ������ʤ����������Ǥ��ɤߤ䤹���褦��ɮ�Ԥ������Ѥ���ɽ���ʸ����ˤ˽�ѹ�������ĺ������
�����������ʸ�Ȥ��Ƥ��뤬���������ƤΥ��ꥸ�ʥ��1909ǯ�Υϡ�������Τ���ʸ�Ǥ��뤳�Ȥ�ǧ�����Ƥ����������������ᡢ����͡�����������������������ʸ����������Ƥ��롣ɮ�Ԥ��ٶ������Ȼפ������������ʿ�ʪ�ˤ�����ֿ��ΰ����˴ؤ�����ʸ�����˾��ʤ����Ȥϻ��¤Ǥ�������ɮ�Ԥ���ؤθ��漼�ˤ����������դΰ�ƣ�������顢�־��衢�������äƾ��ʤ�ȯ���Ƥ����Τʤ顢�����Ҥ��餤�ͤ��Ƹ��ۤ�褦�ˤ��ʤ����פȤ�����ʸ��Ƴ���ä���ΤǤ��롣����ů�ؤ����פʤΤǤ������������ޤǤ⤳�줫����⤹�����Ƥϰ�ĤΥ�ǥ븽�ݤȤ�������Τ�˾�ޤ����Ȼפ����������ä�����Ǥ���С�C.
�ɡ��������� �����쥢��������Ĥβֿ��ΰ������狼�äƤ����ΤǤ��롣
�ﴱ���ȿ����˦�������Ρ�����ҤȤϡ�
���ϡ�������Τϡأ�,��Ϲȿ����Ǥ�����ʪ���١��أ�,���̵��������ʪ����꿧�Ǥ��������뵡���٤Ȳ��⤵��Ƥ�����������Ϥ����ޤǤ⣱�Ĥιͤ����ʳ�ǰ�ˤǤ��ꡢ������η����Ǥ��뤳�Ȥ�ǧ����������ï������Ҥ��Τ�Τ�ȯ�����Ƴ�ǧ�����櫓�ǤϤʤ����������Ƥΰ����Ҥ��ɤ��������ξ�ˤ���Τ��狼��ʤ��Τ������Ȥꤢ���������Ҥ�̾����á��Ҥ�̿̾���Ƥ���ΤǤ��롣�Ȥˤ������Τ褦�˹ͤ���Ȥ��ޤ������ͼ��������Ǥ��뤫��Ǥ��롣
�����Τ褦�˰���Ūñ�̤Ȥ��Ƥθ�ŵŪ�ʡȰ����ҡɤ������ξ����������졢��˦ʬ�����̤���̼��˦�˵�§Ū����ã����뤳�Ȥ�����Ƥ��������Τ褦�ʰ����Ҥ˻��ۤ��������ι�ư��ˡ§�����ηϤŤ����Τ����ط��������ء٤Ȥ������ΤǤ��롣
�����θ�1953ǯ��ȥ���ȥ���å��ˤ��ģΣ�����Ť餻��¤��ȯ���ϡ������Ҥ���������Τι�¤��������äȤʤꡢ�����Ҥ�ʪ��Ū���Τϱ�����������ӥ�����3��ʬ����ʤ�˻��Ǥ��ꡢ�ģΣ���2�ܤΥݥ�̥��쥪���ɺ����餻����ˤͤ��줿��¤���Ĥ��Ȥ����餫�ˤ��줿��
���ģΣ��ʥǥ�������ܳ˻��ˤϤ��礦�ɲ桹�����ڤ�Ͽ������ơ��פߤ����ʤ�Τǡ��ơ��פ��Ǻ�����ȥ����Ǥ��롣���Υơ��פˣ��Ĥ�ʸ�������ʥ��ǥ˥�ˡ��ԡʥ��ߥ�ˡ��ǡʥ����˥�ˡ��áʥ��ȥ���ˤ��¤�Ǥ��롣�����Ͽʹ֤�ޤư��ʪ����ʪ���Ƥ˶��̤���ʸ���Ǥ��롣�ʹ֤�30���ФĤޤ�60���Ĥ�ʸ���ǤǤ����������ġ����ʤߤ˥����������륹�Ϥ鷺��9000�Ĥ�ʸ��������ưפ˿�����Ƥ��ޤ������Ͽʹ֤�10�ܰʾ��ʸ����200��800���Ф�ʸ����Ȥä������̤��ű�ʲ֤�餫���Ƥ��롣�����Фδֳ֤�0,34����ʥʥΥ�ȥ롢10�������ˤʤΤǡ�����˦������ģΣ�������Ĺ��27,2���ȿ������ʤ��褦��Ĺ���ˤʤäƤ���ΤǤ���ʿʹ֤��κ�˦�ˤΣģΣ�������Ĺ��2���ˡ���������������Ĺ����Ф��������������̿�ΤʤΤ��Ȥ����С�ñ��ˤ����Ȥϸ����ʤ����ģΣ�����̵��٤����Ǥ���ȡ�����α�������������Ҥ�¿���ޤޤ�Ƥ��ꡢ������ȿ������ε�ǽ�����餫�ˤʤäƤ��ʤ���
������¾�������ΣģΣ��٤�إߥȥ���ɥꥢ�ģΣ��٤ʤɤ�¸�ߤ��Ƥ��ꡢ��˦�������˴�Ϳ���Ƥ���������ʤɤΡإ��ǥ�������Ρ٤ȡ������������Ρ٤⤢�ꡢ��ǽ�����ΤǤʤ��ģΣ����ۤȤ�ɤǤ��롣
�����ǥꥺ��ˤ���������Ҥγ�ǰ�Ϸ���ȯ���δ���Ūñ�̡��Ĥޤ�ʣ������ҡ��������ˤȤ���Ƥ��������θ塢���Ǥ���䤬������ȼ�äơʣ������ҡ������ǡˤȹͤ�����褦�ˤʤä������ߤǤϺǾ��ΰ�������ˤ����������ʪ���ϥ��ߥλ��Ǥ��뤳�Ȥ��Τ��Ƥ��롣
���ʾ塢����ȡ���ʪ�Τ��������Τϡ��شﴱ�٤Ǥ��ꡢ�ﴱ��Х�Х�ˤ���ȡ��ȿ��٤ˤʤꡢ�����ȿ��ϡغ�˦�٤����ޤäƤǤ��Ƥ��롣��˦����ˤϡغ�˦�ˡ٤�¸�ߤ������κ�˦�ˤ���ˡ������Ρ٤����롣
�����������Τ���ˤ�27�����Ĺ���Υơ��פ���Ť餻��������äƤ��롣���Υơ���˰�������4�Ĥ�ʸ����1600��ʸ����800���С˹�߹��ޤ�Ƥ���ΤǤ��롣����ʸ���������Ĥ����ޤꡢ���줫��Ҥ٤褦�Ȥ�������ҤȸƤ�Ǥ����Τˤ��ƤϤ�뤳�Ȥ��Ǥ��롣
�����ʤߤ˿�ʪ��˦����¹�˼����Ϥ����������̤�桹���ɤ��ʪ�˴�������ȡ�1000�ڡ�������ʤ�ɴ�ʼ�ŵ��7700��ʬ���������뤳�Ȥˤʤ롣����7700��ʬ�ΰ��������ƺ�˦�κ�˦�ˤ���¸����Ƥ���Τ����顢���ब�ե��åԡ��ǥ������˾������¸���Ƥ����٥�ʤɤޤ��ޤ��������Υ�٥�ˤ�ã���Ƥ��ʤ��ΤǤ��롣
������Ϥޤ��äϡ����鲿ɴ���������ᤫ�β��ڡ���ʬ���������뤫ʬ����ʤ��������ȥ��ι���βֿ��˴ؤ�������ͼ��ˤĤ��ƤξϤǤ��롣
���βֿ�����ꤹ������
�����ۼ¸���������θ����饰�롼���̤�ʬ����줿�����Ĥ��Υ��ȥ�丶����Τ��˼�������ɽ���ˡ�ɽ����ˤϣ��/�ңҤȣã�/���ȼ�����Ƥ��뤬������ϣ���ΰ����ҷ���ɽ���Ƥ��ꡢ�ä���ӣ�Ϲȿ����Ǥ�����ʪ����Chromogen�������⥲��ˤ����ä�ͥ���ǣ㤬���������Ҥ���ӣ��̵���Υ����⥲���ͭ���ο��Ǥ�ž�����뵡�������ʤ������ʪ����꿧�Ǥ��������뵡����ɽ�����Ҥ�ͥ�����������������äƹȿ��βֿ��ˤʤ뤿��ˤϣäȣҤĤޤ�����ʪ���ȿ�������������¸�ߤ�Ʊ����ɬ�פǤ��롣�㤨�УҤΰ����Ҥ������Ǥ���У��Ȥ��ä�ͥ�������Ҥ�¸�ߤ��Ƥ���֤ˤʤäƤ��ޤ��ΤǤ��롣�����ȹ礻���դξ��Ǥ�Ʊ�����Ȥ������롣�Ĥޤ�ä����������ҤǤ���㤬¸�ߤ�������ǡ��Ҥ�ͥ���Ȥ��Ƥ�ȿ��βֿ��������ʤ��ΤǤ��롣
ɽ��������ΰ����ҷ����Ѥ������ȥ�丶��ʥ���Сˤ�ʬ��
| ���롼����ccRR�� | ���롼����CCrr�� |
| C.gaskelliana��var.alba C.intermedia����Alba�� C.labiata��Alba�� C.labiata����Harefield��Hall�� C.loddigesii��Alba�� C.loddigesii��Stanley�� C.lueddemanniana��Alba�� C.mossiae��var.wageneri C.��obrieniana��Alba�� C.skinneri��Alba�� C.speciosissima��Alba�� C.trianaei��Alba�� C.trianaei��Broomhills�� C.trianaei��Verdonck��s�� C.warneri��Alba�� |
C.eldorado��Alba�� C.harrisoniana��var.alba C.mendelii��Alba�� C.percivaliana��Alba�� C.schroederae��Alba�� C.trianaei��Alba�� C.warneri��Alba�� C.warscewiczii ������������Firmin��lambeau�� |
���롼�ף���ǤϤɤθ���Ʊ�Τ������碌�Ƥ⤹�٤���֤����������롼��II��Ǥ�Ʊ�ͤ����Τμ��������ޤ�롣���������롼�ףɤȥ��롼��II�δ֤Ǥθ��Τθ��ۤǤϤ��٤ƹȲ֤�������ΤǤ���ʿޣ��ˡ�
���ǤϤ��������Ʊ�Τθ��ۤ�����줿�Ȳ�Ʊ�Τθ��ۡ��⤷���ϹȲ֤�ե������ʼ��ȼ�ʴ�ˤ�����硢�����κ�˦�����ҷ��ϡ��أã�/�ң�٤�������ʬ�����������˦�ʲ�ʴ����ҡˤϡ��أãҡ١��أã�١��أ�ҡ١��أ��٤Σ������פ��ͤ���졢�����������ޤ�Ƥ���ƣ��ΰ����Ҥ��ȹ礻�Ȥ��Ƥϡ�������16�̤�ˤʤ롣������ǡ����Σ�-/��-�Υ����פ����ȹ礻�����̤�ǹȲ֤�餫���롣����ʳ����ȹ礻�ϣ��̤�Ȥʤꡢ��֤�餫���롣�������äơ��Ȳ֡���֡��������Ψ�Ȥʤ�ʿޣ���ɽ���ˡ�
���������ºݤ���֤ΰ����ҷ��ϣ��̤�ΰۤʤ�����ҷ��ã�/��ã�/����/�ңҡ����/�ң��/���¸�ߤ��롣����飵�Ĥΰۤʤ�����ҷ������Ʊ�Τ���ۤ����Ƥ�Ȳ֤������롣��������֤��������Τϡ��أ��/���������ۥ�ˡ٤ΰ����ҷ����Ĥ�Τ˸¤��Ƥ��롣
���Ĥޤ긽����֤κǿ����ۼ�Ʊ�Τ������碌�Ƥ⡢��֤����ޤ�Ƥ����ݾڤϤʤ��ʤ��ΤǤ��롣ȿ�Ф˹Ȳ֤ΰ����ҷ����ã�/�ңҡ��ã�/�ңã�/�ңҡ��ã�/�ң�Σ��Ĥ��ͤ����뤬���Ȳ֤���֡��Ȳ֤ȹȲ֤���ۤ����Ƥ����٤ϰ㤨�ɡ��Ȳ֤Ф������ޤ�Ƥ��ʤ���ɬ����֤������뤳�ȤȤʤ롣
������
����������ʼ���Ѥ�������������ۤ�Ԥ������Τ褦�ʰ����ͼ���������Ƥ��뤳�Ȥ��Ф��ơ�æ˹���Ƥ��ޤ������������ϡ�������Τϥ��饤�Ȼפ���ɮ�ԤȤ��Ƥζ�̣�ϡأ����ۤ�����μ����Ĥκ������٤ˤ��롣�߳ػ��塢����ʪ�ξ�硢�֣����ۤ�����4000�Ĥ֤���������Ĵ������٤��פȶ���ä����Ȥ����뤬�������ԤǤʤ��¤���ʿ�ʪ�ǤϤʤ��ʤ����ο��̤Ǥ���ȴ�ϤǤ��ʤ���ΤǤ��롣���ߤ��¸���Ƥ�����ָ��Τ��Ѥ��ơ�����������ۤ�Ԥ����롼��ʬ����������ΤǤ��롣

�ޣ� �ۤʤ�����ҷ����ĥ��ȥ�丶�異��Фΰ����ͼ�
ɽ�� ���ƣ��ʹȲ֡�Ʊ�Τθ��ۡʥ���ե��ˤ�
�ƣ�����֤θ�������١���Ω�������ϼ�ɽ��
���Ʊ�Τ���Ȳ֤���
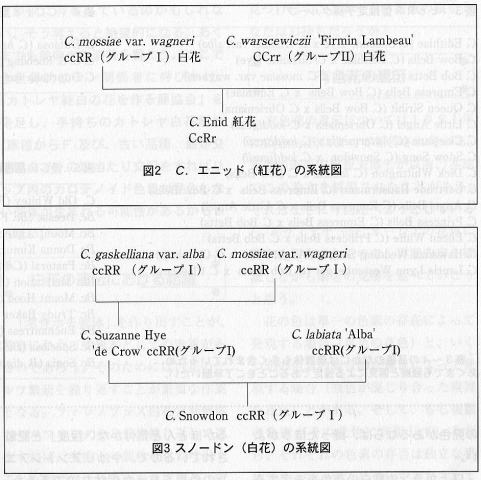
���㤨�н���Ȳ֤Ǥ���C.Enid��
C.mossiae 'Wagneri'(���ң�)��C.gigas 'Firmin
Lambeau'(�ãã��)��Ƥ˻��ĻҤǤ��ꡢ���ܤ˻ĤäƤ���á����˥åɤϥ��ߥ���Х����פΡǥ�ʥ��ƥåɥͥ������ǤȤ������ΤǤ���������˥��ƥե���С�
�ե����쥤�����䥺���ۡ��Τ褦�ʥ��ߥ���Ф��ò֤ߤ�������Τ������̹Ȳ֤�Ĥ���Τ��Ф������ߥ���Ф������Ǥ��ä������������Ƥ����ʼ�Ǥ��ä��褦�������������˥�ʥ��ƥåɥͥ������������ο��������ʣ����Ρˡ������ʣ����Ρˤ���𤵤�Ƥ���Τǡ�ñ��˺���Ρ֣�-/��-�������̤�ˤϤ����ʤ��Τ��⤷��ʤ���
���Ʊ�Τ�����֤���
����������֤ˤʤ���⤢���Ƥ������ʿޣ��ˡ��á����Ρ��ɥ�Υ���ե���������ңҷ������ʼ�ʸ�����ۼ�ˤȤθ�����ޤ줿�Ҥɤ�ϡ�Ʊ�������ҷ����ңҤȤʤꡢ���٤���֤�������Ȥ����Ƥ��롣���μ���Σ��ĤȤ��ơ����������ǽ����ָ��ۼ�Τʤ��ǡ��á��ץ���٥륹�����ңҷ��ȿ���Ǥ��롣�����θ��ﹽ���ϡ�C.gaskelliana
var.alba, C.mossiae var.wagneri, C.trianaei
var.alba�Σ���Ǥ��ꡢC.Edithiae ��C.Suzanne
Hye���ᤷ���ۤ��줿���ᡢ�ãã�η줬������ʤ��ä��餷�������ңҷ��ȳο����뤿��ˤϡ����롼��II�ȸ��뤫����Ҥ��̤ꡢ�ҤϤ��٤ƹȲ֤Ȥʤ�ˡ��������ʼ��ե���������Τ��褤���Ҥ����٤���֤ʤ顢���ңҷ��Ǥ��������������ۤ�ξ����Ǥ⥢��ȥ����˥���졢�֡��ȡ���ȡ��ġˤ�ȯ��������ʤ��,���ä������褤��
| C.Edithiae�������������� (C.Suzanne Hye x C.trianaei var.alba)
C.Bow Bells������������ (C.Edithiae x C.Suzanne Hye) C.Bob Betts (C.Bow Bells x C.mossiae var.wagneri) C.Empress Bells (C.Bow Bells x C.Edithiae) C.Queen Sirihit (C.Bow Bells x C.Obrieniana) C.Little Angel (C.Obrieniana x C.loddigesii) C.Claesiana (C.intermedia x C.loddigesii) C.Snow Song (C.Snowdon x C.loddigesii) C.Dick Whittngton (C.labiata x C.Bow Bells) C.Gertrude Hausermann (C.Empress Bells x C.Bow Bells) C.Angel Bells (C.Empress Bells x C.Little Angel) C.Princess Bells (C.Empress Bells x C.Bob Betts) C.Eileen White (C.Princess Bells x C.Bob Betts) C.Hawaiian Wedding Song (C.Angel Bells x C.Claesiana) C.Laurie Lynn Westenberger (C.Bob Betts x C.Claesiana) |
| ��ɽ���������ʼ�Τʤ��ˤ��ܿ��Τ�¿���ޤޤ�Ƥ��뤿�ᡢ�����ޤǤ����Ǥ��и��ˤ�����Ǥ��뤳�Ȥ�λ���ꤤ������ |
| C.Dubiosa�������������������� (C.harrisoniana alba x C.trianaei
alba) C.C.G.Roebling (C.mendelii alba x C.harrisoniana alba) C.Dupreana var.alba (C.warneri x C.warscewiczii) |
���ʾ夬���ι���βֿ�����ꤹ�����ǤǤ��뤬��������ĺ�����Ǥ������������ңҷ��λ�¹�ϡ����������˻ĤäƤ���ΤǤ������������ңҷ���¹���ȤϤä��ꤤ�����Τ������ˤ��β�ǽ���˶ᤤ��Τ����Ȥ���ȡ���������Ǹ���dz������Ƥ����Τ�C.Princess
Bells�����롣��Ҥ��̤���ңҷ��Ȥ��äƤ褤�����Τ�ʤ���
�����ˣ���������Ƥ����Τˡ�C.Earl��C.Tiffin
Bells�����롣C.Earl�ϣ������οƤˤ�����C.Barbara Billingsley���Ϥä��ꤷ�ʤ��ΤǤ��롣�����ʼ�ϲ���trianaei
var.alba���ᤷ���ۤ��Ƥ��뤬��trianaei var.alba�ϣɡ�IIξ���롼�פ�°���뤿����ңҷ���trianaei
var.alba���Ѥ����Ƥ��������䤬�Ĥ롣C.Barbara Billingsley�� C.Bow Bells�λҶ��Τʤ��Ǥϡ�C.General
Patton'Cheer'AM/JOS�Ȥ������Τ����뤬��������1957ǯ�ˤ�����Ͽ�ˡ֥�å�Ÿ�����˻�Υԥ�ۡ��륹�ݥåȿ����ꤢ�뤬�ۤȤ�ɵ��դ��ʤ����١פȵ��ܤ���Ƥ���Τǡ���Ϥꥢ��ȥ����˥�ο��Ǥ��ä����ΤʤΤǤ�������C.Earl�ϣ�������ˤ���٤����⤷��ʤ���
���ޤ���C.Tiffin
Bells��Ʊ�ͤǣɡ�IIξ���롼�פ�°����C.warneri�������Ѥ����Ƥ��뤿�ᡢ���ǤΤ�����ңҷ��˶ᤤ�ʼ�Ȥ����Ƥ��롣������C.Tiffin
Bells�οƤ�������C.White Blossom��C.Cowaniae��C.Concinnity,C.Intertexta, C.Angelina,
C.Bronacha, C.Mrs.Myra
Peeters�ʤɤˤ⵿�Ǥ������롣
��
�����߾嵭��ξ������Ǥ��ʤ��������ʼ�ϰʲ��Τ褦�Ǥ��롣�ä�Bc.
Deesse�λ�¹�����٤Ʋ�������Bc.Deesse�ϡ��ƣ������dowiana���Ѥ����Ƥ��뤳�Ȥ䡢�ƣ������C.warscewiczii�ʣãã���롼��II�ˤ��Ѥ���졢�ޤ�B.digbyana�η줬���äƤ��뤿�ᡢ����ʣ���ʰ����ҷ��Ƥ���餷����
| C. Old Whitey����������������(F1����ˣãã���Ѥ����Ƥ���� Bc. Deesse (Bc.Ferrieres x C.Lamartine) Bc. Mount Anderson (C.Bow bells x Bc.Deesse) Bc. Donna Kimura (C.Princess Bells x Bc.Mt.Anderson) Bc. Pastoral (C.Mlle.Louise Pauwels x Bc.Deesse) Blc.Meditation (Bc.Deesse x Lc.Fedora) Bc. Mount Hood (Bc.Deesse x C.Claris) Bc. Trudy Baker (Bc.Mount Hood x C.Estelle) Bc. Enchantress (Bc.Empress x C.Fabia) Bc. Speciosa (Bc.Digdyna-mendelii x C.schroederae alba) Bc. Sonia (B.digbyana x C.Hesta alba) |
���ңҷ�����¹���롼����°���뤳�Ȥ��Ǥ��ʤ��ʼ�Ϥ����λ�¹��ͭ���֤�Ĥ����ǽ�������˹⤤���Ȥ�������Ǥ�������
���Ǹ������ȡ������ޤǤ���֤˴ؤ�������Һ¤��ʣá���ˡʣҡ���ˣ�����ΰ����Ҥ�������Ϳ���Ƥ���Ȥ������Ȥ�������Ȥ�����硢�ʲ��Τ褦�ʤ롣
(1)
�����ҷ���Ƚ�Ǥ��뤿��ˤϥ��롼�ףɤ�°���븶��ȥ��롼��II��°���븶��ȼ��������֤Ȥ����碌�뤳�ȤǷ��ꤵ��롣ξ���롼�ףɡ�II�ȸ��Ƥ�Ҷ������٤���֤��餤����硢���ο��ꤵ�������ҷ��ϣ���������ۥ�ˤǤ��롣�����ۥ��Ĥ��뤳�Ȥϡ������ά�塢���˽��פʰ�̣�����Ƥ��롣���������ۥ���Ѥ�����硢���롼�ףɡ����롼��II����鷺�����λ�¹�Ϥ��٤���֤ˤʤ뤿�ᡢ�ֿ��ʳ��ΰ���Ū�Ѱۤβ�ǽ�����뤳�Ȥ��Ǥ��뤫��Ǥ��롣���������ۥ���ꥯ����ˤ����ưݻ����Ƥ����С����פʿƳ��Ȥʤ�Ǥ������ʥ���դˤ���˿��������褤���⤷��ʤ����Ȳ֤ޤʤ��ƤȤ��Ʋֿ������ͼ��ʳ��η�����ʬΥ�����Ѱۤ��٤�������ۥ��Ĥ��뤫��Ǥ���ˡ�
(2)��û��Ƚ�̤�����ˡ�ϼ��������ָ��Ρʸ���Ǥ���ۼ�Ǥ�褤�ˤ�ե��������Ƥߤ뤳�ȤǤ��롣���ξ�硢��֤Ф��꤬�餤���Ȥ��Ƥ⥰�롼�ףɤ����롼��II��°���뤫��ʬ����ʤ���������������ꥹ�Ȥ�Ҥ�Ȥ������Ĥ�Ĵ�٤뤳�Ȥˤ�ꡢ�������ٿ���Ǥ���Ǥ�������
���ʾ�������ϣ����Τ����ˤ����뤳�Ȥǡ������Τ䡢�����Τˤ��������Ѥ��ʤ����Ȥ��Ǥꤷ�Ƥ����ʤ��ƤϤʤ�ʤ�����ҤΣ��ңҷ��Ȥۤ����ꤷ��C.Bow
Bells�Ǥ����⣳���Ρ������Τθ��Τ���ȴ����Ƥ��뤿�ᡢ���ߤ���ָ��Τ��ܿ��ΤǤ��뤳�Ȥ�α�դ��Ƥ����ʤ���Фʤ�ʤ���
����������ȥ����˥�ˤ��ֿ���ȿ���ɽ�����Ƥ����Τϡ�ȯ�������ϰϤ��졢�֡��ȡ��硢�Ĥȹ����������Τ��ƹȿ��ȸƤ�Ǥ����ΤǤ��롣���ߥ���Фθ��Τ⤹�٤ƹȲְ������Ƥ����ΤǤ��롣���ʤ���������Ǥ�֤ΰ����˥���ȥ����˥��Ǥˤ��ȯ��������йȲְ����ʤΤǤ��롣
�����ȥ�����֤Υ�åסʹ��ˤˤϲ������褹���Τ�¿����������å���Υ������Υ��ɡʲ����ο��ǡˤΰ����ͼ����̤�¸�ߤ��Ƥ���Τ��⤷��ʤ��������ͤ������˾Ū�ˤʤ롣�����ޤǤ��������ʻ�����Ӥ�����������������Ū�˥��ط��Ԥ˸Ƴݤ����إ��ȥ�����β֤���������٤�ȯ����������Υ��ȥ����ָ��Ρʸ��狼��ƣ��ڤӡ��Ť��ʼ�ǿ����ۼ�ޤǡˤ�����������ۤ�Х�å���Υ������Υ��ɿ��Ǥ�ȯ�����ʤ����Τ����ޤ���ǽ�������뤫�⤷��ʤ���
�֤ΰ����ˤ��������
���������ۥ���Ρ٤���Ф����Ȥ�������ˤ錄�äƿ��ڤȤ��Ƥΰյ�������ΤǤ����������Τ���ˤ���֤Υ�����˿����֤����Ȥ����פʺ�ȤȤʤ롣�ե���Υץ�����ֱ������ΤΥ���ե����������巫���֤�����ȴ����С������ۥ���ΤĤ��������Ȥ�̴�ǤϤʤ��Ȥ����Ƥ��롣�����ۥ������ȴ��Υ�ꥯ���������������פˤʤ�Τ������θ��Τ���˿���������ǰ��ꤷ��������ԤäƤ�����ΤǤ��롣�֥�ꥯ�����Ĥβ��ʤ��⤤�פȴ������Ƥ������Ϥ��ҤȤ⥻��ե����������巫���֤��Ƥ����٤��Ǥ��������Ϥ����������ۥ�Ĥ��������Ȥ��Ǥ�����ȴ�δ�ʤ��Ϥ���������������
ͭ���֤ΰ���
ͭ���֤ΰ����ˤĤ��Ƥϣ�������ǯ��Fenton��ˤ�ä���𤵤�Ƥ��롣ɮ�Ԥλվ���ƣ������Τ��������ܻ�Ǥ�����ʸ����⤵�줿���Ȥ����롣����Ϥޤ��˺���Ҥ٤褦�Ȥ��Ƥ���C.
�ɡ���������
�����쥢�βֿ��˴ؤ������Ǥ���Τǡ��������Ƥ��ǧ���ʤ���ɮ�Ԥθ������⤷�Ƥ������Ȼפ���
�֤ο���ñ��ο��Ǥ�¸�ߤˤ�ä�ȯ���������ñ���βֿ��ˤȡ������Ĥ��ο��Ǥ�ʣ��Ū��Ư���ˤ�ä�ȯ���������ʣ�����������ä�ʣ���ʲֿ��ˤȤ����롣�����ơ��⤷ʣ���ο��Ǥ����βֿ��˹����Ƥ���ʤ顢���줾��ο��Ǥ�¸�ݤ���Ω�ʻ����Ǥ��äơ��̡��˰�������ȹͤ��Ƥ褤�����Τ褦�ʰ����ͼ���ͤ���Τ����˺�����פ��롣�������äơ��ºݤ���֤Ȥʤ뤿��ˤ�ñ�˹ȿ��ο��Ǥ��礱������Ǥʤ���ͭ���ο��ǤϤ��٤Ƥ��äƤϤʤ�ʤ��Τ��������Ǥ��롣���Ҥ���֤ΰ����ͼ��ϡ����Τˤ����ȡ���֤Ȥ������֥���ȥ����˥��Ǥ�������֡ס��Ĥޤ�֥���ȥ����˥�ϡ�̵���֡פΰ����ͼ��Ȥ������Ȥˤʤ롣�������Υ��ɤ�¸�ߤ��뤿��ˤϲ������뤤�������Ǥ��äƤ⡢�������ե����ޤि���п��Ǥ��äƤ⡢����ȥ����˥��Ǥ��ޤޤ�ʤ���С��������֤ȹͤ��ơ����Ҥ���ɽ�ˤ������äư����ͼ���Ƚ�ǤǤ���ΤǤ��롣
���ȥ�����֤�褯����С���å���˲����ο��Ǥ��ΤäƤ����Τ���������¸�ߤ��뤷���������������п��ʤɤ��٤ƥ���ȥ����˥��Ǥ�¸�ߤ��Ƥ��ʤ���֡ʥ���ȥ����˥�ϡ�̵���֤⤷���ϥ���ȥ����˥�Ͽ��ǡ�����֡ˤ��㤨����ΤǤ��롣
�п��˥���ȥ����˥��Ǥ��Τ�ȡ��ޤ�dz��ζ����碌���褦�ˡ��п����ֿ����㿧�Ȥʤꡢ�����ϤǤϲ������ֿ��������Ȥʤ롣�������֤ο��Ǥ������äƤ��ʤ��������������п������βֿ�������Ǥ����������������ΤǤ���ʲ������п��Ძ�п������ѤǤ������Ĵ�¤Ȥ�����Ρˡ�
���줫�顢ͭ���֤ΰ����ͼ�����⤹�뤬������Ū�˲���ͭ�פʾ����˳���������ä�����⤢�뤫�⤷��ʤ���
���ȥ��Υ���ȥ����˥��ͭ����
���ޤǥ��ȥ�����֤ΰ����ͼ���Ҥ٤�ݤˡ���ְʳ��٤ƹȲ֤�ɽ�����Ƥ��������ȥ�����ɽŪ�ʲֿ���Ȳ֤Ȥ������֥���ȥ����˥��ͭ���֡פ�ɽ������Τ����ΤǤ��롣����ȥ����˥�ˤϿ�Ĵ�ΰۤʤ뤤���Ĥ��μ��ब���äơ��ȿ����濴�Ȥ��ơ��ȿ��˥�������뤤���Ŀ����ä�ä��褦��ȯ���˻��ΤǤ��롣���줬ʣ���ˤޤ�ʤ������ȥ��ξ����Ф��Ƥ���ֿ���̥�ϤǤ�������
�����ǡ������Υ���ȥ����˥�μ������ꤹ������Ҥ�ʬ���äƤ���С����˺ݤ��ƶˤ��ͭ�ѤʤΤǤ��뤬��������������Ϥ���Ƥ��餺����ǰ�ʤ��������Ǥ��ʤ����ᤷ�����ʡ����ȥ�����ز֤���̯����̯�ʿ��礤�˥ҥȤϴ�ư���Ƥ���Τˡ�����İ����ͼ���ʬ���äƤ��ʤ��ΤǤ��롣
���Ǥ�ǻ�٤ˤĤ��Ƥϡ��ޤ������Ĵ�᤹���̤ΰ����Ҥ���ߤ����礬¿���ΤǤ��뤬��������Ϥ���Ϥ���Ƥ��ʤ����ޤ�������ޤǡ��ããңҡ��ããңã�ңҡ��ã�ң�Ȱ����ҷ�����äƤ⤳���٤ƹȲ֤�ɽ�����Ƥ����������ҷ����㤦�ȼ㴳���Ǥ�ǻ�٤��ۤʤ���⤢�뤬�����ΤȤ������ȥ��ǤϤ��Τ褦������𤵤�Ƥ��ʤ��褦�Ǥ��롣
����Ǥϡ��ɡ��������ʤλ�¹�˴ؤ���ֿ��ˤĤ��Ƥ��äˤդ�Ƥ�������
�ɡ��������ʤ��Υ�ӥ��������Ѽ�Ǥ��ä���
Cattleya
labiata���Ѽ�Ǥ���Cattleya labiata var. dowiana�ʰʲ�C.
dowiana�ˤϲ��֤�
���ȥ��Ǥ��뤬�����βֿ��ΰ����ͼ��ϼ㴳Ĵ�٤��Ƥ����Fenton1951�ˡ������Ǥ��ҳ�����ˤ��������Ƥ��������������Ȥˡ��Ρ��ɡ��������ʤϥ�ӥ��������Ѽ�Ȥ���Ƥ������ȤǤ��롣�ɡ��������ʤη����ˤϥ�ӥ������η줬ή��Ƥ���ΤǤ��롣
�������ɼԤ˿֤ͤ��������ȥ�䡦��ӥ������ȥ��ȥ�䡦�ɡ��������ʤȤϡ��ɤΤ褦�ʷ����ΰ㤤�ˤ�äơ���ʤ���ˤΰ㤤��Ƚ�̤��Ƥ���ΤǤ�����������˿֤ͤ�ȡ��֡زַ��٤�زֿ��Ѱۤ����٤�س��ѡ٤�س��ִ��٤�ظ����ϡ٤⤹�٤ưۤʤ�ǤϤʤ�����������郎�㤦�Τ��פ��ⶵ���줽�������������ˤ�������η��������������ǽ�ʬ��ʬ�व��Ƥ��������������ȵ�����äƤ��ޤ��Τ��ݤ�ʤ��������äƼ�ΰ㤤��������Ƥ���ΤǤ���������
��ʪ�ذ��̤Ǥϡ��ָ��ۤ��Ƥ⻨�郎�Ǥ��ʤ�������郎�ۤʤ�פ��������Ƥ���Τ��������ʿ�ʪ���ü�ǡ����⻨��ʥ���ե������֥�Τ��ȡˤ���������ֻ���ʥץ饤�ޥ�Τ��ȡˤΤߤʤ餺��°�ֻ���ʥ�ꥪ���ȥ쥢�ʤɡˤ⤤�Ȥ��ñ�ˤǤ��Ƥ��ޤ�������ϥ��ȳ��Ǥ������ݡʾ��ʤ����������äƤ����Ȥ��ä���ͳ�ˤǤϤ��뤬��¾�ζȳ��ΰ��Ȥ���ϡ����ʿ�ʪ��ʬ�ब�ְ�äƤ����ǤϤʤ������Ȥ�����������褦�����Ĥޤ�ʬ�ष�����Ƥ����ʤ����Ȥ�����Ǥ��롣
Cattleya labiata var. dowiana �βֿ��ΰ���
����ȥ����˥�Ͽ��Ǥ˴ؤ��Ƥϣ�����ɽ�������֥���ȥ����˥��̵���֡פǤ��ꡢ����˲������Ǥ��ä�ä���Τȹͤ����Ƥ��롣�������Ǥ����Ƥ��Τ��Ƥ��ʤ��Τǡ������Ǥ�ñ�˲������ǤȸƤ�Ǥ����������β������Ǥ�ȯ����ʤ�����Ҥϣ٤�ɽ������롣���ΰ����Ҥϥ���ȥ����˥��Ǥξ��Ȥϵդˡ��٣٤�٣��ǤϿ��Ǥ����������������������ۥ�ˤˤʤäƽ��ƿ��Ǥ���������ΤǤ��롣�Ĥޤ겫���֤Ȥ��������������Ǥ��롣
�������ݥ����
���ɡ��������ʤβ����֤η����������Ǥ����
�����ޤǤ�ޤȤ��ȡ�C.
dowiana�ΰ����ҷ��ϣ���������ɽ���Ǥ��롣������Ф��ơ����ȥ����������Ȳ֤ϣããңң٣٤�ɽ�����졢���ˤ��줫����������֤ϣãã��٣١����ңң٣٤�ɽ������ơ��������٣ٰ����Ҥ�ޤ�Ǥ��뤳�Ȥˤʤ롣�������äơ������θ����٥�θ��ۤǤ�C.dowiana�Ȥΰ��廨��Ϥ��٤ư����Ҥ˴ؤ��٣��ʥإƥ��ˤǤ��äơ��������Ǥ���������ʤ����Ĥޤꡢ��֤ȸ��ۤ���Уã���٣����뤤�ϣ��ң�٣�����֤Ǥ��뤷���Ȳ֤ȸ��ۤ���Уã�ң�٣��ǹȲ֤Ǥ��롣
�������ݥ����
���ɡ��������ʤΥץ饤�ޥ�Ϥ��٤Ʋ������Ǥ���������ʤ�����
���ɡ��������ʤ��֤Ȥ���ۤ���Ȥ��٤��֤ˤʤ��
���ɡ��������ʤȹȿ��֤Ȥ���ۤ���Ȥ��٤ƹȿ��֤ˤʤ��
��
��ɮ�Ԥ��Ԥä��ץ饤�ޥ�������C. luteola
���п��֡ˤȤθ��ۤǤϡ��ɡ��������ʤ��������褦�ʡ��ɡ��������ʤ��ä���ʲ����֤����ޤ�Ƥ���Τǡ��쳵�ˤϤ���ˡ§�̤�ˤϤ����ʤ��餷����
���Τ褦�ˡ��������ˤϡ������֤俧Ĵ���Ѳ��������Τϸ���ʤ����Ȥ����������λ���μ��ȸ��ۡʣ٣�
�أ٣��ˤ���ϣ٣١��٣����٣������������줾��ƱΨ������Τǡ��������Ǥ�ޤ��Ρʣ٣٤ȣ٣��ˤȴޤޤʤ���Ρʣ����ˤ����������γ��������뤳�ȤȤʤ롣���λ��郎�Ǥ���С���֤Ȳ��֤��椬�������Ǹ���뤷�����λ��郎�Ȳ֤Ǥ���С��Ȳ֤ȥ�����β֤��������������뤳�ȤȤʤ롣�����ǽ��Ƹ��줿������ϡ��Ȳ֤˲������ä�ä���ΤǤ��ꡢ�����˥�����ο��Ǥ����줿�ΤǤʤ����Ȥϡ����狼�ꤤ��������Ǥ�������
�������ݥ����
���ɡ��������ʤ���¹�ˤϲ����֤����ޤ�Ƥ����Ψ���⤤��
�������Ǥ������˴�Ϳ��������Ҥ�ʣ����
�¤Ϥ⤦����ʣ���ʤΤǤ��롣�����Ǹ��줿���֤䥪����ϡ���֤�Ȳ֤Ǥʤ����ȤϳΤ��ʤΤǤ��뤬�������䥪����Ȥ��äƤ�ǻø�����̤Ǥ���ΤǤ��롣����ϲ������Ǥ�������ʤ�����Ҥ���ĤǤϤʤ����Ȥ���ΤǤ��롣
�ɡ���������
�����쥢�ΰ����ҷ�
C.dowiana�ΰ����ҷ������������������������ɽ������С������Ǥ��ʤ���ʤ������ʤߤ��������ιȲ֤ϣããңң٣��٣��٣��٣��������٥����֤ϣãã��٣��٣��٣��٣����뤤�ϣ��ңң٣��٣��٣��٣��Ǥ��롣�Ĥޤ겫�����Ǥ�������ʤ룲�Ĥ���Ω�ʰ����Ҥ�٣��٣��Ȥ��롣�������äơ�C.dowiana���������ιȲ֤λ������ϡ��ã�ң�٣������٣������Ȥʤ�C.dowiana�ȸ����٥����֤Ȥλ������ϡ��ã���٣������٣��������뤤�ϣ��ң�٣������٣������Ȥʤ롣�����ޤǤϲ������Ǥ��о줷�ʤ��ΤǤ��뤫�顢��̤�ñ�������Ǥ��褦��
ɽ�� �ɡ���������
�����쥢�ȸ����٥����֤ΰ��廨��ʥץ饤�ޥ�ˤ�
�����ҷ��ڤӤ��μ��ȸ��ۡʥ���ե��ˤˤ����벫���֤�ʬΥ��
ɽ���Ϥ��ΰ��廨��μ��ȸ��ۤη�̤��Ƥ��롣�ä��ñ�ˤ��뤿��ˡ�C.dowiana�ȸ����٥����ְ��廨��ˤĤ��ơ����μ��ȸ��ۼ����Ǥβֿ���ʬΥ���ͻҤ�������Ƥ��롣ǻ�������ϣ����������������Ȥʤäƽ��Ƹ����ΤǤ��äơ������������뤤�ϣ�������
�����Ǥ�ø������α�ޤ뤳�Ȥ���ʬ����Ǥ�������
�⤷�����ΰ��廨���C.dowiana���ᤷ���ۤ����ʤ�Фɤ��ʤ�Ǥ������������η�̤⤳��ɽ�������Ǥ��롣C.dowiana�ϣ����������������Ǥ��뤫�顢��ʴ���뤤����ҤΣ�������
����������Ф��η�̤�������뤳�Ȥˤʤ롣��̤ϡ��ʣ٣������٣������ˡ�ø�����ʣ٣��������������ڤӣ��������٣������ˡ������ʣ����������������ˡᣱ���������Ǥ��롣
C.dowiana�ȹȲ֤θ��ۤη�̤���ǫ���ä����顢������Ǥ�³������ۤ�ʣ���Ȥʤ롣���廨��ޤǤϡ�C.dowiana�ϣ������֤Ȥʤ���Ѥ�뤳�Ȥʤ����ҿƤ��������Ȳ֡ʣããңң٣��٣��٣��٣��ˤǤ���Ф��٤ƹȲ֡ʣã�ң�٣������٣������ˡ������ޤ������ʼ﷿�Ȳ֤Ǥ���С����Ҥ������ˤ������äƹȲ֡ʣã�ң�٣������٣������ˤ���֡ʣã���٣������٣�����
�����ң�٣������٣�����
�������٣������٣������ˤ�������˲�������֤������ʤ����Ȥ�������������θ��۷�̤ϡ�����ʣ���Ǥ��äơ���˽Ҥ٤�������ø�����Τۤ����Ȳ֤˲����ξ�ä�������˶ᤤ��Τ䡢�ȿ���ø�����ξ�ä��Ϥ��˥�������ä��ȿ��������Ƥ��롣
�á��ҡ��٣����٣�
���ΰ����Ҥ��ߤ�����Ω�Ǥ��뤳�Ȥ�˺�줺�����Ϥ���С����٤Ƥ��ȹ礻��Ω�ɤ�ɽ��ɬ������Ϥ��Ǥ��롣��äȤ⡢���Σ��Ĥΰ����Ҥ��ߤ�����Ω�Ǥ���Ȥ������Ȥϡ���ʴ���뤤����Ҥΰ����ҷ��ǣ��Σ����16�ˡ��κ�˦�ǣ��Σ����256�ˤΰ����ҷ���¸�ߤ�����Ȥ������ȤǤϤ��롣�ҥȤ�Ƭ�ǤϤ����դ�Ǥ���夲���⤷��ʤ������Ȥϥ���ԥ塼���ˤޤ����褦��
���ȥ�䡦�ɡ��������ʡ������쥢�ڤ���
�ðۤ���������ؤ��åפ��ġ�����ʥ��ȥ��θ���إɡ���������
�����쥢��
������ǥ���̮�ˤ����Ʋ����Ϥΰ����Ҥ�ʲ����������طϥ��ȥ��θ���ϡ��������ꥫ�Ǥϡإɡ��������ʡ١�������ӥ��Ǥϡإ����쥢�١��ڥ롼�Ǥϡإ�å����٤ȸƤФ�Ƥ��롣�����٤ơإ�ӥ������٤��Ѽ�Ȥ��ư����Ƥ�������⤢�ä��������ߤˤ����Ƥ���ۤɤΰ���ʰ�����¸�ߤ��ʤ��Ȥ��äƤ����ǤϤʤ����������إɡ��������ʡ������쥢�٤Ȥ�������Ϥɤ�ʤ�ΤʤΤ���������
���ƹ�Υ��θ��ҤǤ��륦�����ʡ���Τ����Ƥ���褦�ˡ������쥢�ϥɡ��������ʤȰ�äƼ郎���ꤵ��Ƥ��餺����Ϥ�¸�ߤ��Ƥ��ʤ��Ȥ����Ƥ��롣����ʸ���ˤ��Ȳ֤η����ˤ��͡��ʤФ�Ĥ������롣��Τϼ��¸³�Τ���������������餷���ַ����ȯ�����뤿��ˡ���Ҥ�����ݤ��롢���ʤ��ͭ���˿���Ԥ����Ȥ�¿��ʲ��ͤ�����Ǥ������𤷤Ƥ��롣��������Ƭ�ˤ⤳�ξ��ܺ٤˽Ҥ٤��Ƥ���������ǰ�ʤ��鸽�ߤˤ����Ƥ�ï��ͼ¸��Ǥ��Ƥ��ʤ��ΤǤ��롣
�����줫��ɡ��������ʤˤĤ��Ƹ��ڤ��Ƥ���������������ŵŪ��̾��ʸ�����鸦�椷�Ƥ⡢��������٤ξ����̤����ʤ����Ȥ��Ƽϴꤤ������
�����Σ�����ǯ�֡�����ϥɡ��������ʡ������쥢�Υ�åפ�̥�����ƺ��ݤ��֤�����̡�������̤λ��Τ����Ϥ餷�Ƥ��ޤä���������������ǽ�����������ᡢ���Σ�����ǯ�֤�³���ȥ����쥢�λҶ�������Ƥ����ΤǤ��롣����ʿ�������Ϥ����������Ϥ��������¤������åפκƸ��ϤǤ��ʤ��ä��褦�Ǥ��롣�����麣���Ǥϼ����ζ�������������ۼ�ǤϤʤ����ű��̥�Ϥ�������줿��åפ������ʪ�Υ����쥢�������Ƥ��롣�ޤ����ܲȤΥɡ��������ʤ��鸡�ڤ��Ƥ�������
���˥������ꥫ���ɡ���������
����ӥ������Ϥ��緿���ȥ������ز֤�Ĥ��롣��åפ��礭��Ÿ����������������ǻ���Ȼ翧�˲��������������������ΤǤ��롣�ڥ��롦���ѥ�Ϥ���߲����Ϥ˻�ȿ���̮�����ä��ꡢ�ڥ���α�ˤ�ȯ�������ꤹ�롣
�����Τ褦�ʷ��������ʼ���ɾ塢�礯���ȤΤǤ��ʤ����פʸ���Ǥ���Ȥ����Ƥ�����������������ꥹ�ȤǤϤ��Υ������ꥫ���ΡإΡ��ޥ륿���פΥɡ��������ʡ٤ȼ��˽Ҥ٤���Ѽ�Υ����쥢�٤�Ʊ���Ƶ��ܤ��Ƥ������ᡢ��������ǯ�֡����ڤȤ��Ƥɤ��餬�Ѥ����Ƥ����Τ��ĤäƤ��������Ĵ�٤Ƥ⡢���ߤΤȤ�����äƤ��ʤ���������ܲ��ܸ��Υ������ꥫ�Ǥϡ��Ρ��ޥ륿���פΥɡ��������ʤϸ������Ǥ��Ƥ���Ȥ����Ƥ��롣�Ĥޤ�Ρ��ޥ�ʥɡ��������ʤ�������õ���Ƥ⤢�ޤ긫�������ʤ��ΤǤ��롣
���Υ�������ʡ�å��줿̿̾����
���ǽ��ȯ�����줿�Τϣ�������ǯ�ǡ�ͭ̾�ʥ�������å����J.Warscewicz�ˤ��������ꥫ�Ǻν����������ꥹ�Υ�����������ä�������Ǥ����ؤ���ߡ����֤��ʤ��ǸϤ�Ƥ��ޤä�������˥饤����Хå���Τ�����줿�Ϥ��δ���ɸ�ܤ�Ĥ��˥�����������夷�ʤ��ä��������Τ����Τ����줫�Ǥ�̵�������夷�Ƥ���С����������餷�����ϥ饤����Хå���Τˤ���������å���δ�˾�̤ꡢ�ѹ�κ��ݲȤǤ���������ˤ��ʤ��̾�����Ĥ����Ƥ����Ȼפ��롣
���饤����Хå���ΤϤ��Σ���ǯ��Σ�������ǯ��������α��ǯ�嵭�˰ʲ��Τ褦�˵����Ƥ��롣
�֥ɡ��������ʤ���μ긵���Ϥ��ʤ��ä�����ˡ��伫�Ȥ�̿̾��������Ǥ��ʤ��ä����Ȥϻ�ο����ˤ��������ʤ���������
�����θ塢��������ǯ�˥������ꥫ�ǥ��������M.Arce�ˤ���ȯ�����ơ������ꥹ�Υ�����ʡ�������ꡢ���Τ����������ӡ��������Veitch
&
Sons)�ǣ�������ǯ�ν��˽鳫�֤����������ƥ�����ʡ���δ�˾�ˤ��٥��ȥޥ���Τ�̿̾���������μ�̾�Ϥ����γ������Ƥ��饤���ꥹ�ر�����ɥ���Ĺ(Captain
J.Dow)��̾��ǰ���ơ��إɡ���������(Dowiana)�٤�̿̾����ȯɽ���줿��
���裱ȯ���Ԥΰո��Ǥ��ä���������ʤǤϤʤ����ɡ��������ʤ�̿̾���줿�ΤˤϤ��Τ褦�ʤ������Ĥ����ä��ΤǤ��롣
�����֤���ħ
�������������ǡ��Х�֤���������Ĺ�����������ľ�¤Ϻ��磴����Τ�Τ⤢�롣�դ�Ĺ�������������������������������դα�ˤϤ鷺���˥�����¸�ߤ��롣���դ�Ĥ��롣
�����ؤ����֤��ؤ��飶�ؤĤ��롣�ַ¤ϣ���������������֤ϲ������۱�˥ԥ���ȿ���̮��ή�졢��åפ��礭�������Ƹ����Ǥ��ꡢǻ��ȿ��˲�����������롣��������餭��
�����Ϥε���
���������ꥫ�Ǥ�������¦�˸����Ϥ�¸�ߤ��Ƥ����褦����ɸ��ǹ⣱���������ޤǤλ��Ϥ���©��������������ɤ���ޤ��������Ƥ��������ΤۤȤ�ɤ��Ͼ�ޤǤ���������⤤�����������Ƥ����Ȥ����Ƥ��롣�����ϰ��ŵ��Ū��Ǯ�ӱ��ӵ����ǡ����٤Ͼ�ˣ�����������ȹ⤯�����̤Ͻ�ʬ������ۤɤ���Ȥ����Ƥ�����ǯ�ֹ߱��̤ϣ����������������������ˤ�ã���롣�����ϣ���飱����Ǹ�夫�����������ߤ�Ф���ʿ�ѵ����ϣ�������ݤġ������������ϣ�����飴���ʿ�ѵ����ϣ�����餤�Ϥ���褦���������Ϥμ������֤Ǥϣ���飵��˳��֤��Ƥ���褦�Ǥ��롣���ޤˣ�����ˤⳫ�֤������𤵤�Ƥ��롣
���˥�����ӥ����ɡ��������ʡ������쥢
���إɡ��������ʡ٤μ����Ϥ��飱,����������ʾ��Υ��Ƥ��륳����ӥ��ˡإɡ��������ʡ������쥢�٤Ȥ�����Τ����롣������ӥ��α�ݲȤ�ñ�ˡإ����쥢�٤ȸƤ�Ǥ���褦�������μ�ϸ��ϤǤϼ������Ρ��ޥ륿���פ�긵���ǥХ�֤�Ĺ��Ǥ��ꡢ���ۤϽ㲫�������ǹȿ���ޤޤ�����åפβ����̮�ϥ������ꥫ����ꤺ�ä�¿�������������������Τ褦�ˤ��������ǥ����쥢�����äƤ��뤿�ᡢ���ۿƤȤ��Ƥ��Τ����¿���Ȥ��Ƥ����ΤǤ������������������Ϲ�ΰ��̤μ�̣�ȵڤӥ������ȼԤȤ⡢���Υ����쥢�κ��ݰݻ�������ΤǤ��ä��餷�����Ĥޤ����˺��ˤ��������ݺ���ʤΤǤ��롣���ߤǤ⤽����ͳ�ϲ�������Ƥ��ʤ���
ȯ���ε�Ͽ
�����μ�ϣ�������ǯ�˥����ꥹ��ʣǡ��ף�����ˤ�������ӥ���ƥ������������Υե�������ζ��ȯ��������ǯ��˥Хȥ顼����̤˥�����ӥ���ȯ���������٥륮���Υ��ǥ���Τˤ�꣱������ǯ�ˡأá������쥢(C.aurea)�٤Ȥ���̿̾���줿���ᡢ���ߤ⸶���Ϲ�α�ݲȤ�ñ�ˡإ����쥢�٤ȸƤֿͤ�¿���餷�������θ塢�ɡ��������ʤ��Ѽ����Σ��Ĥ������Ȥ������顢����������ꥹ�Ȥˤ�ξ�Ԥζ��̤Ϥ��줺Ʊ���ɡ��������ʤȤ��Ƶ��ܤ���뤳�Ȥˤʤä��ΤǤ��롣
�����Ϥ�ʬ�۾����ڤӴĶ��Ѱ���
������ӥ��ˤ����륪���쥢�θ����Ϥ⥳�����ꥫ�˹������褦�Ǥ��롣�ä˥���ֳ���������ߤ�¿���ߤ���褦�Ǥ��롣�ޤ���¦��̮�������̡�¨����ʿ��¦�γ�ȴ�����������������٤μ���ʥ����˼������Ƥ���Ȥ������⤢�롣������ӥ��Ǥϡ����Υ����쥢���礭�����Ĥ�ʬ�व��Ƥ���褦�Ǥ��롣���Ĥϥ��祳�����ФΥ��롼�פ�ñ��ˡإ��祳�٤ȸƤФ졢�⤦�����Υ��롼�פϥ���ƥ���������������������϶軺�Τ�Τ�ؤ���ñ�ˡإ���С٤ȸƤФ�Ƥ��롣���̤��ܰ¤ϡ����祳�ϲ��������ܤΤߤǤ���Τ��Ф��ơ�����Ф����ܤ�ȯã������åפ����٤����뤯����ˤϤۤȤ�ɲ����쿧�Τ�Τ⤢��Ȥ����Ƥ��롣
�����֤���ħ
�֤ϣ������ؤĤ��롣�ַ£�����������������ۤϲ��������ˤϥΡ��ޥ륿���פΤ褦���ֻ翧���ӤӤ뤳�Ȥ⤢��餷�����إ����쥢�٤Υ�åפ˸��Ф������ܤϺ�Ω�ä������Ǥ��ꡢ��åפϥӥ����ɾ��ǥ٥�٥åȼ���ǻ�ֻ翧��������������;��˲���ζڤ����ꡢ��åפα�ޤǿ�Ÿ���Ƥ��뤳�Ȥ�¿�������β�������ܤ��ۤȤ�ɲ����쿧�˥٥��ξ��֤�ȯŸ������åפα郎�鷺���˹Ȼ翧�κ٤��Ӥ�Ĥ����Τ⤢�롣��Ϥꡢ�إ����쥢�٤��������������ϥ�åפβ����̮��ή����������ͤȥ��ѥ롦�ڥ���ν㲫�����褷������ȥ饹�ȤǤ��롣��������餭��
������ˡ
�����ݤ����Ū����Ǥ�����Τ��餤���Ƥ��롣��ʬ���俢���ؤ���Ԥ�����ϲƤ��Ǥ��ɤ�����Ϥ��Τ褤����ݥ��ȡ��С����ߥå�����˭�٤ʸ��̡��̵������٤ΥХ���ۤɤ褯�ݤ����Ȥ褯���ݤǤ���褦�Ǥ��롣���꤬��Ĺ��ϡ�Ŭ�٤ʿ����Ԥ��Ф褤�Τ��������������鲤������Ϥ�������ڤä������褤�����֤���С������̤�������ɤ�����̲���Ͽ������ˤ��������褤������ʸ���Ǥϡ���������ߤ���ݤ������褤�Ȥ����Ƥ��롣��������ʬ�ʲ��١���������������Ф����˸¤롣
�������ϤǤ��륳����ӥ��μ�̣�Ȥ����ϡ����ɤ�Ĺ�����ڤä������ҡ����ڤ������դ��ƺ��ݤ����ꡢȭ�˿���������ꤷ�Ƥ���褦������������֤�в᤹��ȴ�����������������˹����������������������Τ������ѿ����������˳��礷�Ƥ䤬�ƥХ�֤�ü�ǥݥ�������դ��Ƹϻष�Ƥ��ޤ����Ȥ�¿���褦����
������
���á��������Ȥμ�����أá��ϡ��ǥ�����(C.hardyana)�٤��Τ��Ƥ��롣�ֿ��������ȿ��ޤ��͡��ʤ�Τ����뤬�����ߤǤϤۤȤ�ɸ����ʤ�����å���β������ܤ��Ϥä�����Ω�����ɡ��������ʤ�ꤺ�äȺ��ݤ��䤹���Ȥ����Ƥ��������꤫�ǤϤʤ���
���ȥ�䡦�ɡ���������
�����쥢�Υ���ե����������Ĥ�����
�������飳��ǯ���ˡ����夬�����쥢�λ��Τ���֤�͢�����Ƥ����ΤǤ��뤬���������夯�ݻ�����Τ⺤��ʾ��֤Ǥ��ä���������˿��뤿�ᡢ���٤����ȼ�ʴ�����Τ��������ͤ��Ȥ�ʤ����Ȥ�¿���ä��褦�Ǥ��롣���������κ���̤Ƥ��Ѥ��٤����Ƥ����Τǡ�ɮ�Ԥʤ�˥Х��ƥ����Ȥ��Ƥɤ��ˤ��Ǥ��ʤ���Τ��ȻװƤ��Ƥ����Τ����Ƥ��롣
1990ǯ�������ɮ�Ԥ��ϥ磻��ؤ��鵢��塢���Υ����쥢���˿��˾��Ф����ΤǤ��롣���꤫��Υ������Ϻ�����פ����Τǡ���ɡ��ե饹���Ĥ������ϥ���ե��Ǥ��ä������˾����ʼ�Ҥ����Τ줺�����������˼塹�����ä��Τǡ��ޤ��塹�����Ĥϥե饹����Ǵ����Ƹϻव����������˥ե饹����Ǻ�������Ϥ��뤿�ᡢ�Ŀ�ʪ�Τ�ĺüʬ���ȿ�������ʬ��������Ȥ��ä��������˥�Ԥä����Ĥޤ�ȯ�ꤷ�������Τ�˥�������˥������ᡢ�����Τ������ä��Ĥ���������Ƥ����Ȥ��ä��̾�ǤϹԤ�ʤ�������ˡ��Ԥä���ɮ�Ԥ��������Τ褦���Ĥ�ե����ƤǤ����Ĥ��˥����쥯�Ȥ˺����������Τǡإ���ե����������ġ٤�̿̾������1997ǯ�˼����ζ����Ĥ�����夬�ä���
C.dowiana var. aurea��Stella�ǤΥ���ո���βֿ�
��ǯ���餾�������ȥɡ��������� �����쥢
�ƥ��ƥ�ǤΥ���ե����������Ĥγ��֤���𤵤�Ƥ��롣������ϰʲ��μ̿��Ȥ���ĺ���������ʤ��ˤϥ�ӥ��������ä���ʤ�Τ�ǤƤ��롣���������餫�˳��ִ�����ӥ������Ȥϰ�äƤ����ꡢ���餫�ΰۤʤ��������äƤ���褦�Ǥ��롣�����ؤ�ؤ�Ǥ��ʤ���С��ɡ���������
�����쥢�μ��ȼ�ʴ�Ϥ��٤ƥ����쥢�����פ����ޤ�Ƥ�����������Ƥ��ޤ��Τ������Ǥ������������Ϥ�Ȥ�ȥ���ե��Τ�Τ����顢���Ҥΰ����ͼ��ˤ�äƲ������ޤ�Ƥ��뤫ʬ����ʤ��ΤǤ��롣ɮ�Ԥε����Ǥϡ��ż�塢�鷺����ȯ��������ä��Τǡ�����������������ƥ�����������ΤǤ��뤫�顢�ɤΥե饹���ˤɤ�ʷ������ޤޤ�Ƥ��뤫�ޤǤ��İ����Ƥ��ʤ��ä������������ΤǤ��롣�㤨���ԥ���ȿ������֤��Ƥ⡢����˥���ե�����С�������˥����쥢�����פ��餤�Ƥ���Τ����顢�����쥢�ΰ����Ҥ��äƤ���Τˤ��Ѥ��ʤ����Ťʰ����ʤΤǤ�������
�����Ϥ����Ǥ��롣����λ��Τ���Ǥ�������������Υ�����ĤǤ�������������Υ��֥���ĤǤ����������ۼ�ָ��ۤΥץ饤�ޥ�Ǥ���������ʣ���ʸ��ۼ�Ǥ�������ϥ��֥�åɤǤ�������������ե���³���뤳�Ȥˤ�ꡢ�����ۥ⤬���ޤ���פ�������ΤǤ��롣
 |
 |
| ƣ¼��˧�� Home Page����ŵ | |
| C.dowiana��var.��aurea��'stella'��selfing��clone | C.dowiana��var.��aurea��'stella'��selfing��clone |
�Ѹ����
�������Һ¡����������ξ�ǰ����Ҥ�������֡������Һ¤ϣģΣ����ΰ����Ĺ�����������롣��Ĥΰ����Һ¤DZ����Ф������Ѱۤ����������Ω�����ҤȤʤ롣�ȴ����ϰ����Һ´֤ΤߤǤʤ�����Ĥΰ����Һ���Ǥⵯ���롣
������ʪ����������շ�ϩ�ˤ����ư��ʳ������뤤��ʣ�����ʳ���Фơ����벽��ʪ�ˤʤ����β���ʪ��
���ۥ���ܹ��Ρˡ�����Ʊ���ܹ��ΤȤ⤤�������ܤ��뤤���Ĥ��ΰ����ҤˤĤ��ơ���Ω�ط��ˤ�������ҤΤ��٤Ƥ���ǽŪ���°�Ū��Ʊ��Ǥ����ܹ��Ρ�����Ω�����ҤǤϣ�������ᡢ����Ω�����ҤǤϡ������£¡�������⡢���£¡������ΰ����ҷ����ĸ��Τ������Τ褦�ʾ��֤�ۥ⡢���뤤��Ʊ���Ȥ������ۥ��ܹ��Τϡ������ˤ��ۤʤ�������ĸ��Τ�§�Ȥ���ʬΥ���ʤ����Ȥ���Ƚ�ꤹ�뤳�Ȥ��Ǥ��롣���٤Ƥΰ����ҤˤĤ���Ʊ���ʸ��ΤϽ�ϤǤ��ꡢ�ۥ��ܹ��Τϼ������뤤��Ʊ�ϸ��ۤ��֤���������ä��뤳�Ȥˤ�ä������롣